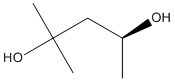
2-methyl-pentane-2,4-diol
Compound of the cristallization buffer (necessary). It is found in multiple place in the structure but found also in active site in a position compatible with substrate
General
Type : Analogue of substrate
Chemical_Nomenclature : (4S)-2-methylpentane-2,4-diol
Canonical SMILES : CC(CC(C)(C)O)O
InChI : InChI=1S\/C6H14O2\/c1-5(7)4-6(2,3)8\/h5,7-8H,4H2,1-3H3\/t5-\/m0\/s1
InChIKey : SVTBMSDMJJWYQN-YFKPBYRVSA-N
Other name(s) : MPD,(4S)-2-Methyl-2,4-pentanediol,(S)-2-Methyl-2,4-pentanediol,(4S)-2-methylpentane-2,4-diol,(S)-(+)-2-Methyl-2,4-pentandiol,(S)-(+)-2-Methyl-2,4-pentanediol,2-methyl-2,4-pentanediol,(MPD)-O4
MW : 118.17
Formula : C6H14O2
CAS_number : 99113-75-4
PubChem : 5288834
UniChem : SVTBMSDMJJWYQN-YFKPBYRVSA-N
IUPHAR :
Wikipedia :
Target
Families : 2-methyl-pentane-2,4-diol ligand of proteins in family: Monoglyceridelipase_lysophospholip || Hydroxynitrile_lyase || ABHD10 || A85-Feruloyl-Esterase
Stucture :
Protein : human-MGLL || manes-hnl || mouse-abhda || bactn-BT4077
References (4)
| Title : Structural insights into the oligomeric effects on catalytic activity of a decameric feruloyl esterase and its application in ferulic acid production - Du_2023_Int.J.Biol.Macromol__126540 |
| Author(s) : Du G , Wang Y , Zhang Y , Yu H , Liu S , Ma X , Cao H , Wei X , Wen B , Li Z , Fan S , Zhou H , Xin F |
| Ref : Int J Biol Macromol , :126540 , 2023 |
| Abstract : Du_2023_Int.J.Biol.Macromol__126540 |
| ESTHER : Du_2023_Int.J.Biol.Macromol__126540 |
| PubMedSearch : Du_2023_Int.J.Biol.Macromol__126540 |
| PubMedID: 37634773 |
| Gene_locus related to this paper: bactn-BT4077 |
| Title : ABHD10 is an S-depalmitoylase affecting redox homeostasis through peroxiredoxin-5 - Cao_2019_Nat.Chem.Biol_15_1232 |
| Author(s) : Cao Y , Qiu T , Kathayat RS , Azizi SA , Thorne AK , Ahn D , Fukata Y , Fukata M , Rice PA , Dickinson BC |
| Ref : Nat Chemical Biology , 15 :1232 , 2019 |
| Abstract : Cao_2019_Nat.Chem.Biol_15_1232 |
| ESTHER : Cao_2019_Nat.Chem.Biol_15_1232 |
| PubMedSearch : Cao_2019_Nat.Chem.Biol_15_1232 |
| PubMedID: 31740833 |
| Gene_locus related to this paper: human-ABHD10 , mouse-abhda |
| Title : Structural basis for human monoglyceride lipase inhibition - Bertrand_2010_J.Mol.Biol_396_663 |
| Author(s) : Bertrand T , Auge F , Houtmann J , Rak A , Vallee F , Mikol V , Berne PF , Michot N , Cheuret D , Hoornaert C , Mathieu M |
| Ref : Journal of Molecular Biology , 396 :663 , 2010 |
| Abstract : Bertrand_2010_J.Mol.Biol_396_663 |
| ESTHER : Bertrand_2010_J.Mol.Biol_396_663 |
| PubMedSearch : Bertrand_2010_J.Mol.Biol_396_663 |
| PubMedID: 19962385 |
| Gene_locus related to this paper: human-MGLL |
| Title : Structure determinants of substrate specificity of hydroxynitrile lyase from Manihot esculenta - Lauble_2002_Protein.Sci_11_65 |
| Author(s) : Lauble H , Miehlich B , Forster S , Kobler C , Wajant H , Effenberger F |
| Ref : Protein Science , 11 :65 , 2002 |
| Abstract : Lauble_2002_Protein.Sci_11_65 |
| ESTHER : Lauble_2002_Protein.Sci_11_65 |
| PubMedSearch : Lauble_2002_Protein.Sci_11_65 |
| PubMedID: 11742123 |
| Gene_locus related to this paper: manes-hnl |