The ESTHER database
The superfamily of alpha/beta hydrolase fold was defined by structural homology. (Follow this link for a list of selected bibliographic references on alpha/beta hydrolases). The three dimensional structures of the lipases from the fungus Geotrichum candidum (GCL) and human pancreas as well as the structure of acetylcholinesterase revealed a very similar topology of the central beta-sheet and the same location of two (Ser,His) of the three residues comprising the catalytic triad.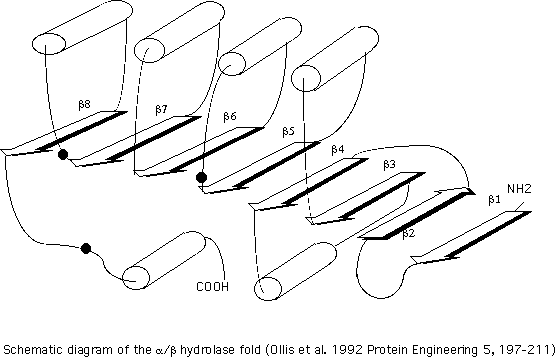 Although these proteins show little sequence homology, the conservation of the topology suggests that these proteins diverged from a common ancestor (Ollis et al. 1992). The figure on the left (click on the icon) is a schematic diagram of the common features of the structure. It consists of a central 5 to 11 stranded beta-sheet in which most strands are parallel (example of order of strands are 12435678 or 21345). This sheet is flanked on both sides by alpha-helices. The core of the structure can be represented as follows:
Although these proteins show little sequence homology, the conservation of the topology suggests that these proteins diverged from a common ancestor (Ollis et al. 1992). The figure on the left (click on the icon) is a schematic diagram of the common features of the structure. It consists of a central 5 to 11 stranded beta-sheet in which most strands are parallel (example of order of strands are 12435678 or 21345). This sheet is flanked on both sides by alpha-helices. The core of the structure can be represented as follows:

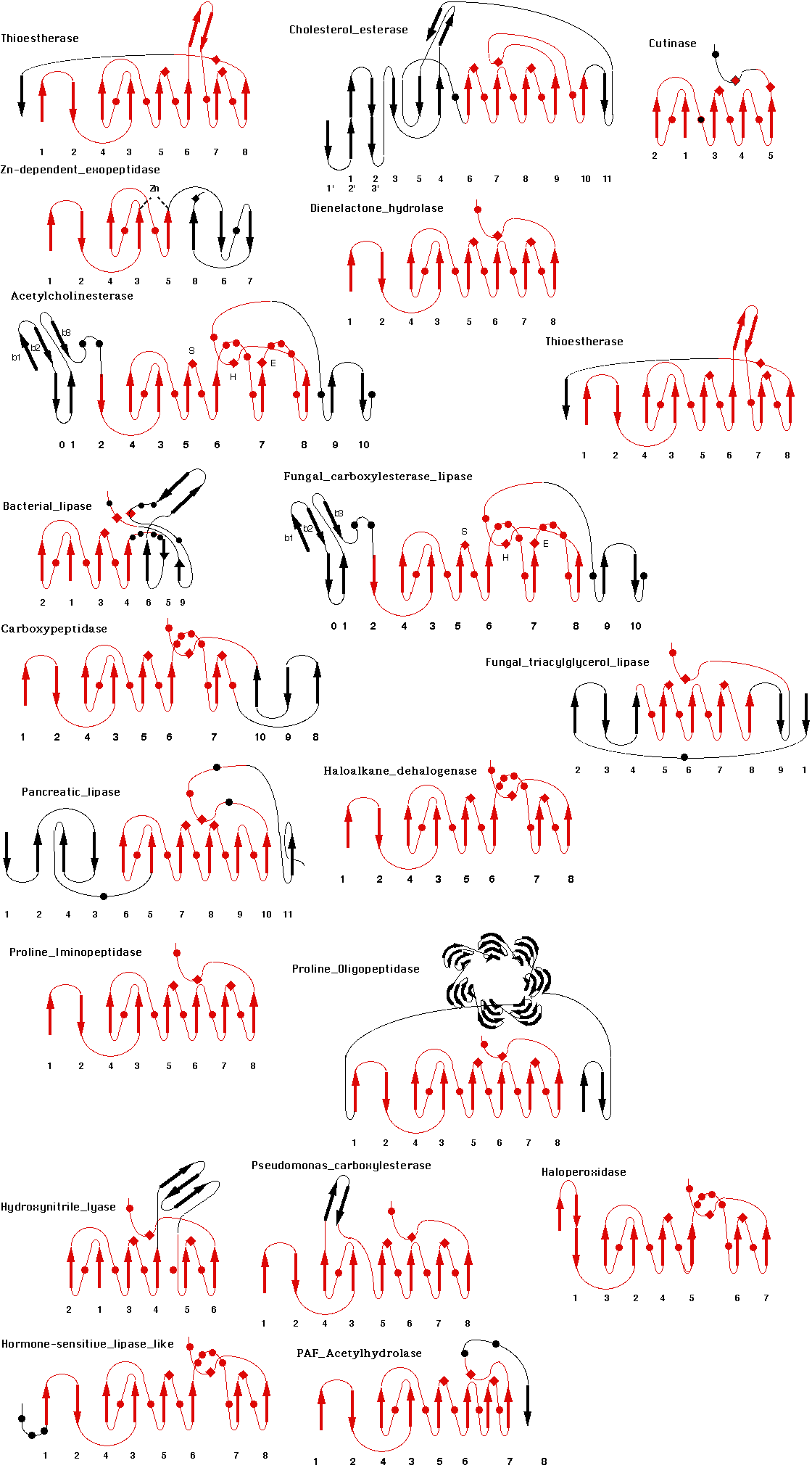
Now many more proteins were shown to belong also to the superfamily. These include carboxypeptidase II from wheat, dienelactone hydrolase from Pseudomonas, haloalkane dehalogenase from Xanthobacter, cutinase from Fusarium solani,Vibrio harveyi thioesterase, Xanthomonas campestris proline iminopeptidase etc.... The superfamily also now includes many peptidases, and epoxide hydrolases. Now Interpro .AB_hydrolase and Superfamily .SSF53474 cover most alpha/beta hydrolases
There are 248 subfamilies as seen in the list of Tables or the table overallTable (old synthese) Now at least 734 different proteins have been crystallized and shown to belong to the superfamily (view liste) . There are 143 families in which at least one protein structure is known. There are 3084 files in the PDB (Protein DataBase ) containing coordinates of native enzymes, or complexes with inhibitors or models. Links to those files are included in the structure chapter of ESTHER or (view liste) .
The diagrams of these have been grouped in a figure: Figure of all
Note A number of structure some of which have been mentionned here as being part of the family, indeed have another fold: the Serine esterase from Streptomyces scabies (1esc), the platelet-activaing-factor acetylhydrolase from Bos taurus (1wab) and rhamnogalacturonan acetylesterase from Aspergillus aculeatus (1deo),the haemagluttinin esterase fusion glycoprotein from Influenza virus C (1flc)
{kind=link}
Our mistake was brought to our attention by Dr. Derewenda and Dr. Mølgaard (see reference Molgaard, Kauppinen & Larsen (2000) Structure, 8:373-383) A name for this family was proposed SGNH-hydrolase. These structures are included in the SCOP database under the Flavodoxin-like fold, the Esterase/acetylhydrolase super family This may include now also the GDSL lipolytic enzymes (for example E. Coli thioesterase/protease I) and the PFAM family PF00657 GDSL-like Lipase/Acylhydrolase
Note The family Zn-dependent_exopeptidase had been mentionned as having some similarities with alpha/beta hydrolase fold family in the paper Ollis et al. 1992. Now it seems they should not be included in the alpha/beta hydrolase fold family. This Zn-dependent_exopeptidase family might be mentionned as "in-laws" together with the SGNH-hydrolase family and the chemotaxis protein CheB. We acknowledge and would like to follow the classifications of experts in the field Heikinheimo P, Goldman A, Jeffries C, Ollis DL (1999)Of barn owls and bankers: a lush variety of alpha/beta hydrolases Structure Fold Des 7:R141-6
Note A fold : (Flavodoxin-like) groups now SGNH hydrolase superfamily Class I glutamine amidotransferase-like superfamily (containing DJ-1/PfpI family), and many others... These are close to alpha beta hydrolases and some time referred as such in papers but are not included in ESTHER
Note Some families group carbohydrates esterase classified as families 1,3,5,7,10 in the CAZy (Carbohydrate-Active enZYmes server )
Note Some families contain lipolytic enzymes from bacteria that have been classified originally by Arpigny and Jaeger in 1999 and updated by Jaeger and Eggert in 2002, Hausmann and Jaeger (2010), Kovacic, Krauss and Jaeger (2019). Correspondance between the classification is found in the following table: Arpigny_Jaeger table. Note that Family II and Family VIII in this classification are not alpha/beta hydrolases but SGNH hydrolases and beta lactamases respectively and are not represented in ESTHER
No other database than ESTHER holds all alpha/beta hydrolase fold proteins together:Interpro, Prosite, Pfam, have multiple entries for subsets of this structural superfamily. A table overallTable shows the correspondance between these database entries and the subfamilies in ESTHER. Now Interpro .AB_hydrolase and Superfamily .SSF53474 fulfill this task also.
The ESTHER old table is now a little to big to be usefull. Each file contains one of the 66581 non redundant proteins/genes. The tables grouped in the overallTable (old: family table, syntheses table) or the structure table may be more usefull.
The Gene_locus nomenclature for these non-redundant entries is a name with 5 characters for the organisms (3 for genera, 2 for the species, except when a common 5 character name exists. ex: ratno is for Rattus norvegicus and human for man. This allows us to keep close to the Swiss-Prot nomenclature). The last characters define the protein, ex: human-ACHE represents human acetylcholinesterase.
During evolution stuctural similarity is preserved much longer than sequence similarity.
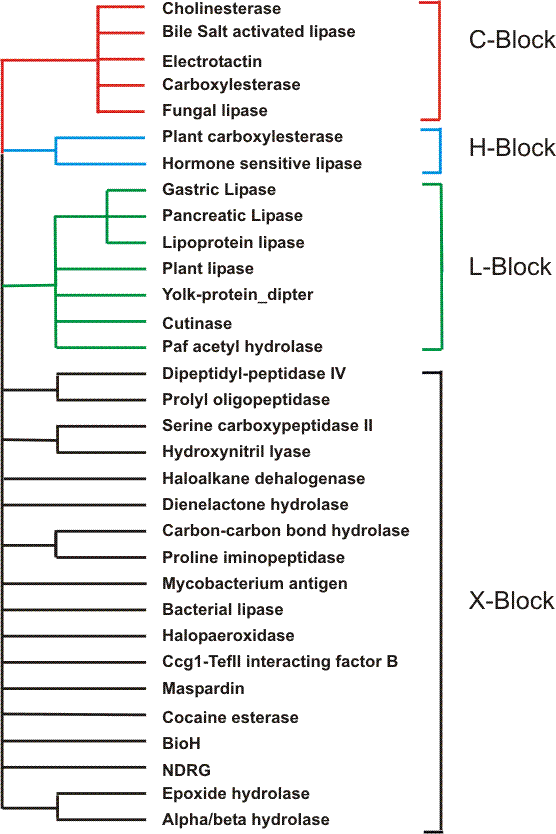
Although the server ESTHER presents information on all alpha/beta hydrolase fold proteins it focuses on a smaller group of proteins of the superfamily for which sequence homology can be detected. This group of sequences includes cholinesterases, carboxylesterases, lipases, and some non enzymatic domains of other proteins (Krejci et al. 1991; Gentry and Doctor 1991; Cousin et al., 1996).A schematic drawing of the relationship of these protein subfamilies follows. (click on the tree icon)
The sequence homologies of these subtypes suggests that they derive from a common ancestor. Although some sequences show little homology, some blocks are conserved.These BLOCKS were used to select sequences for the ESTHER database.On the BLOCK page you will find the definitions and the prototypes of the sequences which have the PROSITE "signature" (PSOO122) of carboxylesterase/cholinesterase called here the
This confirms the identity of this H subfamily.
Different families of proteases also belong to the alpha/beta fold structural family. (See the peptidase liste, and the Merops database) These are until now all included in the SC clan of families of the Serine-type peptidases.For the moment, families S9 (except S9D?) S10 S15 S28 S33 are alpha/beta hydrolases
The S9 family (Prolyl oligopeptidase, Dipeptidyl-peptidase IV, Acylaminoacyl-peptidase) has a PDOCfile PDOC00587, the S10 family (Carboxypeptidase C) has a PDOCfile PDOC00122, the S15 family (Lactococcus X-Pro dipeptidyl-peptidase), the S28 family (Lysosomal Pro-X carboxypeptidase)