Acetonecyanohydrin
General
Type : Cyanide
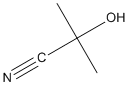
Chemical_Nomenclature : 2-hydroxy-2-methylpropanenitrile
Canonical SMILES : CC(C)(C#N)O
InChI : InChI=1S\/C4H7NO\/c1-4(2,6)3-5\/h6H,1-2H3
InChIKey : MWFMGBPGAXYFAR-UHFFFAOYSA-N
Other name(s) : 2-Hydroxy-2-methylpropanenitrile || 2-Hydroxyisobutyronitrile || 2-Methyllactonitrile || Acetone cyanhydrin
MW : 85.10
Formula : C4H7NO
CAS_number : 75-86-5
PubChem : 6406
UniChem : MWFMGBPGAXYFAR-UHFFFAOYSA-N
Wikipedia : Acetone_cyanohydrin
Target
Families : Hydroxynitrile_lyase
References (3)
| Title : The targets of acetone cyanohydrin neurotoxicity in the rat are not the ones expected in an animal model of konzo - Soler-Martin_2010_Neurotoxicol.Teratol_32_289 |
| Author(s) : Soler-Martin C , Riera J , Seoane A , Cutillas B , Ambrosio S , Boadas-Vaello P , Llorens J |
| Ref : Neurotoxicology & Teratology , 32 :289 , 2010 |
| Abstract : |
| PubMedSearch : Soler-Martin_2010_Neurotoxicol.Teratol_32_289 |
| PubMedID: 19932169 |
| Title : Reaction mechanism of hydroxynitrile lyases of the alpha\/beta-hydrolase superfamily: the three-dimensional structure of the transient enzyme-substrate complex certifies the crucial role of LYS236 - Gruber_2004_J.Biol.Chem_279_20501 |
| Author(s) : Gruber K , Gartler G , Krammer B , Schwab H , Kratky C |
| Ref : Journal of Biological Chemistry , 279 :20501 , 2004 |
| Abstract : |
| PubMedSearch : Gruber_2004_J.Biol.Chem_279_20501 |
| PubMedID: 14998991 |
| Gene_locus related to this paper: hevbr-hnl |
| Title : Structure determinants of substrate specificity of hydroxynitrile lyase from Manihot esculenta - Lauble_2002_Protein.Sci_11_65 |
| Author(s) : Lauble H , Miehlich B , Forster S , Kobler C , Wajant H , Effenberger F |
| Ref : Protein Science , 11 :65 , 2002 |
| Abstract : |
| PubMedSearch : Lauble_2002_Protein.Sci_11_65 |
| PubMedID: 11742123 |
| Gene_locus related to this paper: manes-hnl |